Имея представление о том, что такое аллели, легко объяснить первые законы генетики — три закона Менделя.
Впрочем, самим Грегором Менделем они были выведены во второй половине прошлого века чисто эмпирически на основе результатов проведенных им скрещиваний растений.
Первый закон - так называемый закон единообразия первого поколения при скрещивании чистых форм. Используем в качестве примера скрещивания таких партнеров: кота и кошку, различающихся по окрасу: кот черный, а кошка - сиамского, или гималайского окраса - сил-пойнт. Причем в обозримом прошлом все их предки имели тот же окрас, то есть оба партнера являются гомозиготными по аллелям интересующего нас гена. Все потомки от такого скрещивания будут одинаково черными. Почему?
Сделаем небольшое отступление. Генетическая информация, заложенная в организме, называется генотипом, а ее внешнее проявление - фенотипом. В конкретных случаях часто говорят о генотипе, имея в виду не весь геном, но какой-то отдельный ген или группу генов. Фенотип определяется генотипом прежде всего за счет взаимоотношения аллелей, входящих в состав первого. Каждый обнаруживаемый у животных или человека ген получает свое название, а для обозначения аллелей этого гена используют одну-две начальные буквы его английского наименования (например, White -доминантного белого окраса, чей аллель может быть обозначен как W). Сиамский, или гималайский окрас - это вариант неполного альбинизма (акромеланизма), вызванный мутацией в локусе гена Colour.
Один из аллелей этого гена обеспечивает равномерный полностью развитый окрас, и, как показывают результаты скрещивания, его проявление у потомства преобладает. Такой аллель называется доминантным, и его символ изображается прописной буквой С. Другой аллель - сиамского окраса - является рецессивным (подавляемым) по отношению к С и обозначается строчной буквой "с" с надстрочным символом “s”.
Итак, родительские особи были гомозиготные по разным аллелям локуса Colour: кот - СС, а кошка - сs сs(рис.5). Каждый из них может давать только один тип половых клеток (в отношении данного гена, разумеется): сперматозоиды, несущие С, и яйцеклетки, несущие сs. При объединении этих гамет в новый организм возможен только один вариант генетического набора Ссs то есть все котята будут и гетерозиготные, и единообразны внешне.
Рис.5 Перераспределение аллелей при смене поколений
Взаимодействие аллелей по типу доминантность - рецессивность наиболее распространено. На уровне механизма действия генов это явление можно объяснить различной активностью образующихся белков (особенно регуляторных): более активные формы будут иметь внешнее выражение, а менее активные или неактивные вовсе не проявятся. Часто аллели, определяющие последние типы белков, называют гипоморфами или аморфами. Если же активность белков примерно равна, можно встретиться с явлением промежуточного наследования. В том же локусе Colour существует еще один аллельный вариант – сb определяющий в гомозиготе другой акромеланистический окрас — бирманский, с незначительным осветлением корпуса и золотистыми глазами. В гетерозиготе с аллелям С (аллелей одного гена у одного организма только пара) сb ведет себя как рецессивный, а вот в сочетании c сs обуславливает промежуточный, тонкинский окрас — более светлый, чем бурма, но темнее сиамского, и специфический зелено-голубой цвет глаз.
Тем не менее при скрещивании гомозигот cb cb и cs cs потомство будет опять-таки однородно по окрасу и генетически гетерозиготно - cb cs.
Вернемся, однако, к черным котятам, полученным от скрещивания черного кота и сиамской кошки. Если скрестить таких потомков между собой, то во втором поколении, при преобладании черных, появятся и котята сиамского окраса. При достаточно большой выборке можно заметить, что первых примерно втрое больше, чем вторых.
При скрещивании гибридов первого поколения между собой в потомстве второго поколения наблюдается закономерное расщепление по определяемому этим геном признаку. Это и есть второй закон Менделя, или закон расщепления.
В самом деле, каждый из гетерозиготных потомков образует два типа гамет: несущие аллель С или аллель cs. При их объединении могут возникать различные комбинации: две подобные родительским — Ссs, и оба исходных гомозиготных сочетания – СС и cs cs. Итак, генетическое расщепление во втором поколении выглядит как 1:2:1, хотя внешне и выражается как 3:1. При промежуточном наследовании генетическая основа и внешнее ее проявление будут - одинаковы - 1:2:1. Для иллюстрации третьего закона Менделя используем для исходного скрещивания животных того же окраса, однако предположим, что у черного кота шерсть короткая, а у кошки — длинная. Длина шерсти определяется геном локуса Long; его доминантный аллель L определяет короткую, а рецессивный l - длинную шерсть. Опять-таки примем, что родители гомозиготны по исходным генам: генотип кота можно записать как CC LL, генотип кошки - cscs ll. Гены, определяющие эти признаки, относятся к разным локусам, а это значит, что у каждого из родителей будут образовываться половые клетки только одного типа (рис.6).

Рис.6 Перераспределение аллелей,
определяющих разные признаки при смене поколений
Все котята первого поколения пойдут в отца — такие же черные и короткошерстные. И гетерозиготные по обоим генам – Ccs Ll. А вот котята второго поколения (от скрещивания этих потомков исходной пары) будут уже четырех типов: больше всего короткошерстных черных, примерно поровну — короткошерстных сил-пойнтов и длинношерстных черных, и совсем мало - длинношерстных сил-пойнтов. Если просчитать значительное количество таких потомков (более 64), то получится примерно такое соотношение: 9 черных короткошерстных: 3 черных длинношерстных : 3 сиамских короткошерстных : 1 сиамский длинношерстный. То есть для каждого признака: окраса и длины шерсти соблюдается то же соотношение - 3 : 1 (12 черных : 4 сиамских, 12 короткошерстных : 4 длинношерстных). Получается, что признаки наследуются как бы независимо друг от друга.
Третий закон Менделя звучит так: аллели каждого гена распределяются в потомстве независимо от аллелей другого гена.
Почему это происходит? Дважды гетерозиготные особи первого поколения могут образовывать четыре типа половых клеток: CL Cl csL csl, а объединяются эти гаметы при оплодотворении равновероятным и случайным образом. Чтобы было проще рассчитать варианты, возникающие при таком объединений, пользуются так называемой решеткой, или таблицей Пеннета, в которой по горизонтали откладываются все варианты гамет одного из родителей, а по вертикали - другого (в рассматриваемом случае гаметы родителей одинаковых типов). На пересечении в каждой клетке записывается образующийся при данной комбинации половых клеток генотип потомка, а на его основе уже определяется фенотип (табл.1).
Таблица 1.
Решетка пеннета для случая скрещивания черного короткошерстного кота
и длинношерстной кошки сил-пойнт
А что получится, если скрестить того же черного кота с кошкой голубого сиамского окраса (блю-пойнт)? Вроде бы обе характеристики относятся к одному и тому же признаку - окрасу. В первом поколении, понятно, все потомки останутся черными. А вот при скрещивании их между собой, кроме ожидаемых черных и блю-пойнтов, появятся и чисто-голубые, и сил-пойнты. При этом распределение окрасов у этих котят будет соответствовать третьему закону Менделя. Получается, что голубой окрас наследуется независимо от сиамского? Это действительно так. Голубой - осветленный - окрас возникает в результате действия рецессивного аллеля d из совсем другого локуса - Dilution, никакого отношения к локусу Colour не имеющего.
Аллель d в гомозиготной форме вызывает осветление любого окраса - черного в голубой, красного в кремовый, сил-пойнта- в блю-пойнт...
Доминантный аллель D обеспечивает интенсивный окрас. Так что генотипы родительских особей следует записывать как CC DD (для кота) и cscs dd (для кошки), а их потомков - как двойных гетерозигот: Ccs Dd (рис.7).
Рис.7 Перераспределение аллелей разных локусов,
определяющих признак окраса
Соответственно может быть составлена и решетка Пеннета (табл.2).
Таблица 2.
Решетка Пеннета для случая скрещивания черного кота и кошки блю-пойнт
Стало быть, такой "признак", как окрас, в реальности определяется не геном одного локуса, а многими генами, рассматривать которые можно только в парных сочетаниях (белый - цветной, пегий - без белых пятен, интенсивный - осветленный).
Однако, если бы вся наследственность подчинялась только менделевским законам, вся племенная работа свелась бы к элемент тарным арифметическим подсчетам. Как говорят генетики, Менделю в: свое время очень повезло, что он случайно начал работу со столь; просто наследуемыми генами. Уже упоминавшееся ограничение двух; из этих законов — то, что они справедливы только при больших выборках.
 |
|
Рис.8 Кроссинговер |
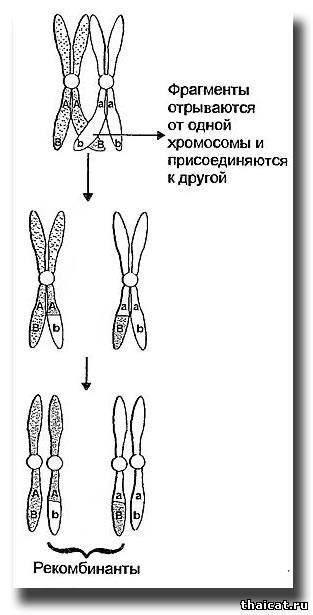
Признание генетиками менделевских законов и попытки применить их к различным организмам показали, что существуют и более значительные ограничения. Причем первым было обнаружено отклонение от третьего закона — так называемое сцепление генов. В самом деле, если при дигибридном скрещивании гены, по котором гомозиготны родители, находятся на разных парных хромосомах, в процессе перераспределения (формирования половых клеток и оплодотворении) эти Пары хромосом как бы независимы друг от друга. Ну а если оба гена расположены на одной хромосоме? Их аллели будут наследоваться вместе, то есть сцеплено - и никакого "независимого наследования" признаков в потомстве второго поколения не будет, а будут наблюдаться исключительно или преобладать только родительские комбинации признаков. Причем чем ближе друг к другу расположены локусы таких генов, тем меньше потомков будет нести отклонения от родительских пар признаков. Казалось бы, таких "отклоняющихся" потомков вообще не должно быть. Однако в процессе деления при образовании половых клеток есть стадия, на которой гомологичные хромосомы обмениваются участками. Чем больше расстояние на хромосоме между двумя локусами генов, тем больше вероятность, что между ними произойдет такой обмен – кроссинговер (рис.8).
Гены, находящиеся на значительном удалении друг от друга, вообще ведут себя как не сцепленные, и наоборот, тесно сцепленные гены обычно наследуются как единое целое. Основываясь на частоте появления в потомстве "отклоняющихся" особей, можно рассчитать, насколько далеко друг от друга расположены на хромосоме те или иные гены. Таким образом составляют так называемые генетические карты разных видов животных. У кошек к настоящему времени мало изучены группы сцепления генов (во всяком случае, тех, которые в первую очередь интересуют заводчиков). Однако, работая с новыми мутациями, список которых постоянно пополняется, рано или поздно фелинолог может столкнуться с этим явлением.
Другой случай отклонения от законов Менделя тоже связан со сцеплением: так называемым сцеплением с полом.
Попробуем скрестить черного кота с красной кошкой.(Разумеется, как и в предыдущих случаях, будем считать, что оба родителя гомозиготны по интересующему нас гену красного окраса, не принимая в расчет другие гены; фактически альтернативной парой признаков будет: красный - не красный (условно - черный)).
Никакого единообразия в первом поколении не наблюдается: все котики - красные, все кошечки - черепаховые, черно-красные. Если поставить обратное (реципрокное) скрещивание: красный кот и черная кошка, то все котята-самцы будут черные, а самки - опять-таки черепаховые (рис.9).
Рис.9 Сцепление с полом: наследование признака красного окраса
Дело в том, что локус Orange, доминантный аллель которого О определяет красный окрас, расположен на половой хромосоме X. А как мы уже говорили, две Х-хромосомы имеются только у самок; (геномный набор 18 пар аутосом + XX), тогда как у самцов Х - хромосома только одна, а вместо второй присутствует Y- хромосома(18 пар + XY). Y- хромосома по сравнению с X очень мала, и большая часть локусов, характерных для X, на ней отсутствует. Однако именно Y определяет мужской путь развития организма. В процессах деления при образовании половых клеток X и Y ведут себя как гомологи, хотя реально таковыми не являются. В результате такой неполной гомологии между ними у самок образуется "избыток" генетического материала по сравнению с самцами. Чтобы такую несправедливость компенсировать, одна из самочьих X -хромосом переходит в неактивное состояние - принимает такую форму, что генетическая информация с нее не считывается. Которая именно из двух Х - хромосом в каждой конкретной клетке зародыша и в какой именно период перейдет в нерабочее состояние - заранее неизвестно, это процесс случайный. (Во всяком случае так считалось до недавнего времени. В последние годы появились работы, показывающие неслучайность инактивации Х - хромосом, или их избирательную, зависящую от генотипа самки, инактивацию).
Все потомство такой клетки (то есть новообразующиеся клетки и ткани) будет сохранять только одну исходную работающую Х-хромосому. На Y-хромосоме локус гена красного окраса отсутствует, следовательно, окрас кота будет определять генотип единственной Х-хромосомы. Какой бы аллель она не несла — доминантный О или рецессивный о - он обязательно проявится, и кот будет полностью красным или черным. У кошек же, гетерозиготных по этому гену – Оо - те клетки, где активна несущая доминантный аллель хромосома, будут вырабатывать только желтый пигмент (определяющий красные и кремовые окрасы), а клетки с рецессивным аллелем – черный. (Точнее и черный и желтый. Подробнее этот механизм будет рассмотрен в разделе о частной генетике кошки.) Это и образует черепаховый, или мозаичный, окрас самок.
Из всего сказанного ясно, что черепаховые коты, которые, хотя и редко (1 на 10000), но встречаются, должны быть мутантами. Выше уже говорилось о котах с тройным набором половых хромосом – XXY. При таком генотипе Х - хромосомы ведут себя так же, как и у кошек, и если эти “Х” - хромосомы будут нести разные аллели гена красного окраса, то и эффект окажется тем же. Однако такой избыток X - хромосом и вызванный им мозаицизм обычно приводят к стерильности кота. Крайне редко попадаются и плодовитые черепаховые коты. В этом случае их окрас тоже вызван мутацией, но происшедшей не в половых клетках родителей, а в одной из эмбриональных клеток. Соматические клетки, то есть клетки тела, мутируют так же, как и половые. Мутацией, приводящей к черепаховости у кота, может быть, например, делеция (утеря) фрагмента Х - хромосомы, содержащего локус с аллелем О. Все пигментные клетки, которые разовьются из такой клетки-мутанта, не будут препятствовать образованию черного пигмента. Остальные же, с неизмененной Х-хромосомой, будут производить только желтый пигмент. Такое сочетание и даст черепахового плодовитого кота. Однако соматические мутации не передаются по наследству, то есть в потомстве такого кота будут или красные, или чёрные самцы. В клетках зародыша при нарушениях процесса деления клеток могут происходить и мутации с изменением числа хромосом - при нарушении процессов клеточного деления. В этом случае у кота образуются клетки и с нормальным набором XY, и с мутантным XXY. Если такая обратная мутация (то есть уменьшение числа X-хромосом) произойдет у кота-трисомика, то он может оказаться плодовитым.
Из характера поведения X - и Y- хромосом как гомологов при образовании половых клеток следует, что теоретически соотношение полов в потомстве должно быть 1:1. Пол будущего котенка определяется только в момент оплодотворения. Однако вполне вероятны отклонения от нормы в потомстве отдельных пар животных и даже некоторых пород (абиссинская).
Во-первых, недостаток самцов в помете может быть связан с их эмбриональной гибелью - гетерогаметный (то есть образующий половые клетки двух типов) пол является более поражаемым.
Это происходит потому, что вредные рецессивные мутации, локализованные на X - хромосоме, обязательно проявятся у самцов в полном их выражении. Во-вторых, возможно существование генетических систем, обеспечивающих преимущественное оплодотворение X или Y-несущими сперматозоидами. Правда, существование таких систем конкретно у кошек пока не показано. Не исключено, что на Преимущественное оплодотворение спермиями того или иного типа, то есть на их активность, можно каким-то образом влиять искусственно (с помощью гормонов, например). Но реально обоснованных практических рекомендаций по этому вопросу пока не имеется.
Признаки, сцепленные с полом, не следует путать с признаками, проявление которых ограничено полом. Гены, определяющие ряд вторичных половых признаков или, например, материнский инстинкт могут находиться не на половых хромосомах, а на аутосомах. "Включаются", то есть становятся активными, они только по мере прохождения организмом физиологического пути развития, определяемого в первую очередь набором половых хромосом.
Долгое время существовало устойчивое убеждение - один ген определяет один признак. Так ли это? Попробуем провести другой эксперимент: скрестить белую голубоглазую и глухую кошку с цветным (хотя бы тем же черным) желтоглазым и нормально слышащим котом. Похоже на тригибридное наследование - как будто имеются три различных признака. Все котята от этого скрещивания будут белыми, » вот во втором поколении появятся и цветные котята. Но что любопытно, среди этих котят голубоглазые и глухие будут встречаться только среди белых потомков. Мало того, среди них могут оказаться и голубоглазые на один глаз (или глухие на одно ухо). И ничего подобного не обнаружится среди цветных котят. Оказывается, доминантный аллель W гена белого окраса определяет не только окрас, но и глухоту, и цвет глаз. Это явление - влияние одного гена на несколько признаков - называется плейотропией. Кстати, расщепление среди белых котят на голубоглазых, разноглазых и желтоглазых, глухих и нормально: слышащих будет отнюдь не менделеевским. (Фелинолог может столкнуться и с другим нарушением менделевского расщепления, по своей природе сходным с плейотропией. Так, при скрещивании двух гетерозиготных по мутации Manx, то есть бесхвостых, кошек, в потомстве на двух бесхвостых будет приходиться один хвостатый котенок (среднестатистически, конечно). Аллель, определяющий бесхвостость, доминантный, значит, не хватает части бесхвостых котят. Оказывается, эта мутация, доминантная в отношении бесхвостости, одновременно летальна в гомозиготном состоянии, то есть ведет себя как рецессивная: зародыши с генотипом MM погибают на ранних стадиях развития).
Потомки второго поколения от скрещивания белой кошки и черного кота не случайно были названы "цветными" - они вовсе не обязательно будут черными. Тот же аллель W подавляет (точнее сказать, маскирует) проявление ряда генов из других локусов, какими бы аллелями они не были представлены. Генетически черные или голубые, тигровые или мраморные - все они внешне будут белыми, если несут аллель W. Такое явление - подавление аллелем одного гена проявления аллелей других генов - называется эпистаз. Эпистаз может быть не только доминантный, но и рецессивный. Например, аллель h гена Hair в гомозиготном состоянии вызывает облысение. Кошки генотипа hh будут голыми, независимо от того, несут ли они гены короткой (L-), длинной (ll), волнистой (rr) или проволочной (Wh-) шерсти.
Итак, один ген может определять не один, а несколько признаков. Самое же главное ограничение законов Менделя - они верны только для моногенных признаков, то есть определяемых одним геном. А' таких признаков очень и очень мало. И вообще, что считать признаком? Ведь получается, что с генетической точки зрения это понятие весьма условное. Тот же окрас можно, конечно, считать признаком, но никак не моногенным. Конечно, его можно - и это гораздо удобнее - рассматривать как пары менделирующих признаков, комбинация которых даст представление об итоговом окрасе. Но отнюдь не все признаки такому "разложению" поддаются - они-то и являются в истинном смысле полигенными, то есть определяются множеством генов.
Для абсолютного большинства характеристик животного, определяемых как "признаки", почти невозможно вычленить отдельные локусы или, во всяком случае, определить действие отдельных генов по внешним показателям. Рассмотрим один из количественных признаков - например, длину хвоста.
Предположим, что итоговая длина определяется генами неких локусов X, Y, Z. При этом гомозиготы XX YY ZZ будут иметь наиболее короткие хвосты, гомозиготы xx yy zz - самые длинные. Те кошки, у которых одна пара генов будет находиться в доминантном гомозиготном состоянии, а другие - в рецессивном, будут показывать промежуточную длину хвоста (Xx yy zz, xx YY zz и т.д.).
Что же касается гетерозигот, то проявление у них признака будет зависеть от степени доминирования аллелей каждой пары. При полном доминировании гетерозиготы Xx Yy Zz будут неотличимы от XX YY ZZ, при неполном - опять-таки покажут промежуточные характеристики.
Мало того, гены полигенного комплекса могут отличаться разной. активностью. Например, гены Х-локуса оказывают большее влияние на длину хвоста, чем гены локуса Z. Тогда особи хх уу ZZ окажутся более длиннохвостыми, чем XX yy zz. Так в крайне упрощенном виде можно представить тип аддитивной, или кумулятивной, полигении. Есть и другие ее типы, например, комплементарная. Признак, определяемый таким образом, будет выражен внешне только в случае, когда каждый из генов полигенного комплекса находится в определенном аллельном состоянии. То есть у кошки генотипа UU Yy zz некий признак выражен, а у имеющих прочие варианты - uu YY zz, UU yy Zz, UU Yy ZZ - нет.
Строго говоря, таких признаков, за которые отвечал бы один и только один ген, практически нет. Даже на те признаки, которые, казалось бы, имеют явно моногенную природу, оказывает влияние; генетический фон — гены других локусов.
Отчасти вариации в проявлении наследственных признаков объясняются влиянием окружающей среды. Наследуется ведь не просто признак в полном его выражении, а, скорее, возможность выражения признака в некоторых пределах, или, иначе говоря, норма реакции. Однако выраженность признака определяется не только условиями жизни организма. Например, степень ослабления окраса, вызванная рецессивным аллелем гена Dilution dd, будет косвенно зависеть от генов, отвечающих за текстуру шерсти. Такие гены, влияющие на проявления аллелей других генов, называют модификаторами.
Для того чтобы охарактеризовать их влияние на проявление того или иного признака, пользуются понятиями экспрессивности и пенетрантности. Экспрессивность - это степень проявления признака. Если она сильно зависит от действия модификаторов, говорят о вариабильной экспрессивности гена, если же нет — о высокой или, стабильной. Пенетрантность определяют как часть особей, внешне проявляющих признак, от числа всех особей, несущих аллели, определяющие этот признак. Пенетрантность может быть полной -например, все кошки, несущие аллель L, имеют короткую шерсть; или неполной - из числа кошек, несущих ген полидактилии (избыточного количества пальцев) Pd, проявляют эту аномалию меньше половины. При значительной выборке пенетрантность может быть вычислена в процентах. Для известного гена доминантного белого окраса White пенетрантность в отношении окраса почти 100%, голубоглазости - около 70% и глухоты - около 40%. Кстати, и экспрессивность, и пенетрантность не являются совсем уж неизменными характеристиками генов. Они значительно варьируют в каждой конкретной кошачьей популяции.














